
Abstrak
- Padang rumput semi-alami dikenal sebagai habitat penting bagi lebah di Fennoscandia. Padang rumput ini dipelihara oleh domba yang berkeliaran bebas, tetapi tidak jelas intensitas domba mana yang paling bermanfaat bagi lebah.
- Kami akan membandingkan kekayaan dan kelimpahan spesies lebah dengan model campuran Bayesian pada berbagai tingkat gangguan oleh domba untuk menguji Hipotesis Gangguan Menengah, sambil mempertimbangkan variabel habitat penting lainnya: jarak ke hutan, ukuran padang rumput, tinggi rerumputan, dan ketersediaan bunga, serasah, dan tanah gundul.
- Implikasi Praktis: Hasil penelitian ini akan memberikan informasi berharga bagi pengelola lahan dalam menetapkan batasan jumlah domba liar yang dilepaskan di padang rumput semi-alami dalam lanskap boreal konifer untuk memberi manfaat bagi konservasi lebah.
1. PENDAHULUAN
Di seluruh Fennoscandia, padang rumput semi-alami menawarkan sumber daya bunga dan tempat bersarang yang melimpah dan beragam (Eriksson et al., 1995 ; Hansson & Fogelfors, 2000 ; Milberg et al., 2020 ; Öckinger & Smith, 2006 ; Pöyry et al., 2006 ) untuk lebah ( Bombus spp.). Namun, karena penurunan pengelolaan ternak tradisional bersamaan dengan konversi ke penggunaan alternatif, area yang kaya spesies ini sekarang menurun (Bele et al., 2024 ; Cousins, 2009 ; Eriksson, 2022 ; Olsson et al., 2000 ; Wilson et al., 2012 ). Norwegia mengalami penurunan 49% pada padang rumput semi-alami dari tahun 1960 hingga 2015 (Aune et al., 2018 ) setelah puncaknya dalam aktivitas pertanian musim panas pada tahun 1850-an (Stensgaard, 2017 ). Di negara tetangga Swedia, Cousins ( 2009 ) menggambarkan penurunan 91% pada padang rumput semi-alami selama 200 tahun. Beberapa studi dari Fennoscandia yang membandingkan padang rumput semi-alami yang dikelola dan ditinggalkan menunjukkan bahwa keanekaragaman spesies tanaman lebih rendah setelah ditinggalkan (Cousins, 2009 ; Johansen et al., 2019 ; Wehn et al., 2017 ), yang dapat memiliki implikasi penting bagi lebah yang juga menurun di seluruh dunia (Goulson et al., 2008 ). Sangat penting untuk memahami tidak hanya apakah habitat ini memerlukan pengelolaan aktif untuk konservasi tanaman dan lebah secara umum, tetapi juga jenis dan intensitasnya secara khusus.
Hansson dan Fogelfors ( 2000 ) menemukan bahwa penggembalaan ternak dan pemotongan rumput lebih efektif dalam mempertahankan keanekaragaman spesies tanaman di padang rumput semi-alami di Swedia selatan daripada pembakaran yang direncanakan dan penebangan semak belukar yang ditargetkan, tetapi dalam meta-analisis global, Tälle et al. ( 2016 ) menyatakan bahwa penggembalaan umumnya lebih bermanfaat bagi keanekaragaman hayati tanaman dan penyerbuk daripada pemotongan rumput. Meta-analisis ini menemukan bahwa penggembalaan yang mengungguli pemotongan rumput untuk tujuan konservasi terbukti khususnya di lokasi Eropa utara dan tengah dan untuk padang rumput semi-kering dan semi-alami (Tälle et al., 2016 ). Di antara para penggembala, Milberg et al. ( 2020 ) dan Öckinger et al. ( 2006 ) menemukan bahwa sapi ( Bos taurus ) dan kuda ( Equus caballus ) lebih bermanfaat bagi penyerbuk dan keanekaragaman tanaman daripada domba ( Ovis aries ) dan kambing ( Capra aegagrus hircus ) di Swedia. Namun Demeaux et al. ( 2024 ) tidak menemukan perbedaan yang jelas dalam kekayaan spesies tanaman dan kelimpahan nektar di lokasi yang digembalakan oleh domba dibandingkan dengan kambing dan sapi di Norwegia barat dengan elevasi rendah. Domba memiliki distribusi global dan sejauh ini merupakan spesies ternak yang paling melimpah di Norwegia (Statistik Norwegia, 2025 ). Dalam jangka panjang, penggembalaan domba mengurangi tinggi rerumputan dan membantu mempertahankan ukuran padang rumput keseluruhan dari padang rumput semi-alami. Petak habitat yang lebih besar sering kali mendukung kekayaan spesies tanaman yang lebih besar, yang pada gilirannya menghasilkan kekayaan dan kelimpahan spesies lebah yang lebih tinggi (Gerner & Sargent, 2022 ; Neumüller et al., 2021 ; Williams et al., 2015 ), tetapi dalam musim tertentu, domba juga mengurangi kelimpahan bunga yang sebenarnya disukai oleh lebah (Scohier et al., 2012 ). Pencabutan bunga secara langsung selama penggembalaan kemungkinan merupakan alasan Milberg et al. ( 2016 ) melaporkan respon positif oleh lebah terhadap ketinggian rumput yang lebih tinggi.
Dinamika yang kontras dari pemeliharaan habitat dalam jangka panjang, dan pengurangan langsung bunga dalam jangka pendek, dapat mengakibatkan beberapa tingkat gangguan menengah yang paling bermanfaat bagi lebah. Banyak sistem terestrial tampaknya memiliki keanekaragaman hayati yang lebih tinggi pada tingkat frekuensi dan/atau intensitas gangguan menengah (Moi et al., 2020 ), namun penerapan umum Hipotesis Gangguan Menengah (IDH) telah diperdebatkan (Fox, 2013 ; Mackey & Currie, 2001 ). Lázaro et al. ( 2016 ) mengamati kelimpahan dan keanekaragaman penyerbuk maksimum pada tingkat penggembalaan sedang pada domba dan kambing untuk beberapa, tetapi tidak semua, putaran pengambilan sampel sepanjang musim di semak belukar Mediterania dan di hutan yang didominasi daun gugur di Swedia selatan. Glimskär et al. ( 2018 ) menemukan kondisi yang paling menguntungkan bagi lebah berada pada tekanan penggembalaan menengah. Pengamatan ini membuat kami menduga bahwa kami mungkin menemukan hubungan ini dalam lanskap boreal konifera Norwegia. Namun, hasil yang beragam dari penyelidikan sebelumnya menunjukkan diperlukannya penyelidikan lebih lanjut.
Beberapa studi padang rumput semi-alami Fennoscandia meneliti hubungan antara intensitas penggembalaan, penyerbuk, dan sumber daya bunga yang mereka andalkan; namun, studi-studi ini mempertimbangkan ternak selain domba (Pöyry et al., 2006 ; Sjödin, 2007 ; Sjödin et al., 2008 ; Söderström et al., 2001 ) atau tidak memisahkan penggembala berdasarkan spesies (Berg et al., 2019 ; Glimskär et al., 2018 ; Milberg et al., 2016 ; Pöyry et al., 2006 ). Sjödin ( 2007 ) hanya mempelajari waktu penggembalaan musiman (Sjödin, 2007 ), bukan intensitas, sementara kategori intensitas Sjödin et al. ( 2008 ) dan Berg et al. ( 2019 ) terlalu luas untuk memeriksa prediksi IDH. Di Finlandia, Pöyry et al. ( 2006 ) memang menunjukkan kekayaan spesies puncak untuk tanaman, kupu-kupu, dan ngengat pada ketinggian vegetasi menengah (indikator intensitas penggembalaan), tetapi penekanan mereka adalah pada serangga sebagai herbivora, dan mereka tidak melaporkan jumlah hewan penggembalaan. Dengan demikian, informasi lebih lanjut tentang intensitas penggembalaan domba tertentu diperlukan bagi pengelola lahan untuk membuat keputusan seputar penetapan batas jumlah domba liar yang dilepaskan di padang rumput semi-alami Norwegia dengan mempertimbangkan konservasi lebah. Mengatasi kesenjangan informasi ini adalah tujuan dari penelitian kami.
Kami menduga pengaruh penggembalaan domba terhadap sumber daya mencari makan merupakan penentu yang lebih kuat terhadap kekayaan dan kelimpahan spesies lebah dalam sistem kami daripada karakteristik yang mendorong ketersediaan lokasi bersarang, tetapi ada faktor lain yang penting untuk mencari makan dan bersarang yang tidak boleh diabaikan. Jumlah tanah gundul dan bahan tanaman mati (‘serasah’) penting untuk bersarang (Potts et al., 2005 ; Svensson et al., 2000 ) dan juga dapat dipengaruhi oleh penggembalaan domba. Kedekatan dengan hutan di lokasi pengambilan sampel tertentu tidak hanya didorong oleh domba, tetapi juga penting untuk lokasi mencari makan dan bersarang (Donkersley, 2019 ; Donkersley et al., 2017 ; Potts et al., 2005 ; Sõber et al., 2020 ).
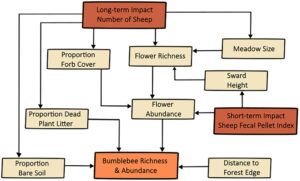
Kami mengaitkan hipotesis kami tentang domba sebagai pendorong kekayaan dan kelimpahan spesies lebah dalam model konseptual (Gambar 1 ). Kami berhipotesis bahwa pada intensitas penggembalaan rendah dan sedang, domba meningkatkan kekayaan spesies bunga dan proporsi forb dengan mengurangi tinggi rumput dan mempertahankan bukaan padang rumput yang besar, tetapi pada intensitas tinggi, rumput mendominasi. Kekayaan spesies bunga yang lebih tinggi menghasilkan lebih banyak bunga yang mekar secara konsisten sepanjang musim dan dengan demikian meningkatkan kelimpahan bunga secara keseluruhan, tetapi tindakan merumput pada periode waktu tertentu memengaruhi jumlah perbungaan yang tersedia dengan cara yang berbeda. Domba mengurangi tinggi rumput dalam jangka panjang, yang pada gilirannya mengurangi jumlah bahan tanaman mati (‘serasah’) yang tersedia untuk bersarang oleh beberapa spesies, tetapi seiring dengan menginjak-injak, meningkatkan tanah gundul yang juga dapat digunakan untuk bersarang. Kami memperkirakan IDH paling baik menggambarkan kontras ini, dengan kurva berbentuk lonceng yang menggambarkan kekayaan dan kelimpahan lebah terbesar pada tingkat domba menengah. Terakhir, jarak ke hutan pada titik tertentu di setiap padang rumput, terlepas dari domba, memengaruhi lebah secara positif, mengingat sumber daya mencari makan dan bersarang tambahan yang disediakan hutan. Semua elemen ini bervariasi di setiap pertanian musim panas kecuali ukuran padang rumput dan jumlah domba yang dilepaskan.
2 BAHAN DAN METODE
2.1 Wilayah Studi
Investigasi ini tidak akan memanipulasi kepadatan domba, tetapi mengambil bentuk eksperimen alami yang memanfaatkan kondisi yang ada. Kami akan mengambil sampel sembilan padang rumput semi-alami (yaitu pertanian musim panas) yang terletak di lereng menghadap timur lembah Sungai Glomma di Daerah Innlandet, Norwegia tenggara (60° LU, 11° BT), pada ketinggian antara 600 dan 700 m di atas permukaan laut (Gambar 2 ). Pertanian musim panas adalah habitat terbuka yang sebagian besar terdiri dari tanaman seperti rumput dan forb. Jenis vegetasi di sekitar pertanian musim panas didominasi oleh cemara ( Picea abies ) di lapisan pohon dan bilberry ( Vaccinium myrtillus L. ) di tanah, meskipun pinus Skotlandia ( Pinus sylvestris ) dan lingonberry ( Vaccinium vitis-idaea ) lazim di daerah yang miskin nutrisi (Larsson & Rekdal, 2000 ). Penebangan kayu di hutan yang keanekaragaman tumbuhannya sudah rendah ini menghasilkan penebangan habis yang didominasi oleh 80% rumput rambut bergelombang ( Avenella flexuosa ; Rekdal & Angelhoff, 2021 ).
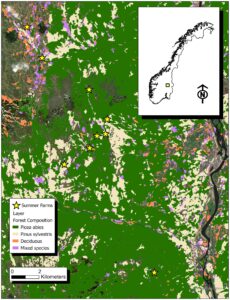
Padang rumput pertanian musim panas berkisar antara 1 hingga 13 ha, tidak dipangkas dan memiliki jumlah domba yang stabil untuk merumput selama beberapa tahun (Tabel 1 ). Domba persilangan Norwegian white dan Old Norwegian Short Tail Landrace (gamalnorsk spæl) dilepaskan pada akhir Mei atau awal Juni dan diambil pada akhir Agustus atau awal September. Domba berkeliaran bebas di daerah tersebut; namun, mereka memiliki kedekatan dengan daerah tempat mereka dilepaskan dan sering mengunjungi padang rumput pertanian musim panas. Perbedaan jumlah domba yang dilepaskan (80–1850, kecuali dua tanpa domba) di pertanian musim panas (Tabel 1 ) memungkinkan perbandingan intensitas yang berbeda untuk menguji IDH. Secara historis, sapi, kuda, dan kambing mungkin juga merumput di pertanian tetapi tidak ada selama >20 tahun. Hewan berkuku besar lainnya termasuk rusa besar ( Alces alces ), rusa merah ( Cervus elaphus atlanticus ) dan rusa roe ( Capreolus capreolus ) jumlahnya dapat diabaikan di padang rumput jika dibandingkan dengan domba.
| pertanian musim panas | Jumlah domba | Jumlah plot dengan perangkap | Jumlah petak dengan jaring tangan | Luas padang rumput (ha) |
|---|---|---|---|---|
| Tempat latihan Skramstad | tahun 1850 | 6 | 3 | 13 |
| Lubang | tahun 1680 | 6 | 3 | 11 |
| Rostadsætra/Blikstadsætra | 610 | 3 | 3 | 3 |
| Klettsætra | 410 | 6 | 3 | 12 |
| Tempat latihan Klovstad | 410 | 1 | 1 | 1 |
| Kota Tingstadætra | 140 | 3 | 3 | 4 |
| Nedre Hemmeldalen | 80 | 4 | 3 | 5 |
| Istana Skyndal | angka 0 | 6 | 3 | 10 |
| Øvre hemmeldalen | angka 0 | 4 | 3 | 3 |
Catatan : Jumlah domba diperkirakan dengan cara mengalikan jumlah induk domba dengan jumlah rata-rata anak domba menurut jenisnya sebagaimana dilaporkan oleh peternak, ditambah jumlah induk domba.
2.2 Periode pengambilan sampel
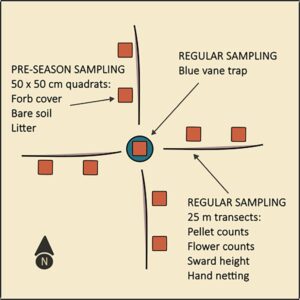
Bahasa Indonesia : Sebelum pelepasan domba pada musim panas 2025 (15 Mei—8 Juni), kami akan memilih enam plot di setiap peternakan musim panas untuk pengamatan berulang, yang mewakili habitat di setiap peternakan musim panas, dari kumpulan titik yang dihasilkan menggunakan alat Buat Titik Acak di ArcGIS Pro 3.0.3 (ESRI, Redlands, CA, AS). Plot akan diberi jarak 100 m dan >10 m dari tepi hutan. Untuk padang rumput kecil yang tidak dapat memuat enam plot, kami akan membuat sebanyak mungkin plot dengan kriteria jarak kami. Selama kunjungan awal, kami akan mengukur forbs, bahan tanaman mati, dan tanah gundul (Bagian 2.5 ; Gambar 3 ), dan membuang pelet feses yang tersisa dari tahun sebelumnya (Bagian 2.6 ). Kami akan menangkap lebah, menghitung bunga, memperkirakan tinggi padang rumput dan melakukan penghitungan pelet sekali pada awal (8–28 Juni), tengah (29 Juni−19 Juli) dan akhir (20 Juli–9 Agustus) musim penggembalaan, dengan jeda minimal 7 hari di antara pengambilan sampel di lokasi tertentu (Bagian 2.3 , 2.6 ).
2.3 Menangkap lebah
Kami akan mengumpulkan lebah di setiap pertanian musim panas dengan menempatkan satu perangkap baling-baling biru (BanfieldBio Inc., Seattle, WA, AS) di tengah setiap petak, digantung 1 m di atas tanah dan diisi dengan 3 dL larutan air sabun. Perangkap akan dipasang selama 24 jam setiap putaran pengambilan sampel dan tidak diaktifkan selama hujan lebat. Setelah diambil, kami akan menyaring isi perangkap dan menyimpannya dalam pusaran air untuk setiap perangkap. Penyortiran dan identifikasi lebah akan dilakukan di laboratorium (Bagian 2.4 ).
Meskipun pengambilan sampel pasif lebah dengan perangkap baling-baling biru umumnya menghasilkan lebih banyak tangkapan, lebih sedikit tangkapan sampingan, dan lebih banyak spesies daripada metode lain dan merupakan metode yang andal untuk memperkirakan kekayaan spesies (Acharya et al., 2022 , 2024 ; Short et al., 2023 ; Stephen & Rao, 2007 ), perangkap baling-baling biru dan metode pengambilan sampel pasif lainnya mungkin tidak dapat diandalkan untuk mengukur kelimpahan dan hubungannya dengan habitat (Kimoto et al., 2012 ; Portman et al., 2020 ; tetapi lihat Bell et al., 2023 ). Memang, Mathis et al. ( 2024 ) mengamati hubungan negatif antara kelimpahan lebah dan kelimpahan bunga menggunakan perangkap baling-baling biru, sementara Kuhlman et al. ( 2021 ), Pei et al. ( 2021 ) dan Westerberg et al. ( 2021 ) melaporkan bias negatif serupa dengan metode pengambilan sampel pasif lainnya (perangkap panci) tetapi mencatat perbedaan menurut musim terbang, warna bunga, dan skala spasial di mana bunga dinilai. Dengan demikian, Mathis et al. ( 2024 ), Briggs et al. ( 2022 ) dan Prendergast et al. ( 2020 ) menyimpulkan pengambilan sampel aktif adalah ukuran yang lebih baik untuk menilai kelimpahan lebah, terutama yang berkaitan dengan habitat. Mengingat kendala logistik dalam penelitian kami, menggunakan baling-baling biru untuk pengambilan sampel pasif akan memungkinkan kami untuk meningkatkan jumlah total plot yang diambil sampelnya; namun, jika kami menggunakan hasil dari perangkap baling-baling biru tidak hanya untuk kekayaan spesies tetapi juga kelimpahan, harus jelas apa yang ditunjukkan oleh jumlah tangkapan yang lebih tinggi dalam plot tertentu. Akankah lebih banyak tangkapan baling-baling biru dalam penelitian kami dihasilkan dari pengambilan sampel di habitat yang kualitasnya relatif buruk karena hanya ada sedikit makanan alternatif dan lebah tertarik pada perangkap karena kurangnya sumber daya lain, seperti yang disarankan oleh Kuhlman et al. ( 2021 ) dan Mathis et al. ( 2024 )? Sebaliknya, lebih banyak tangkapan lebah biru dapat terjadi di habitat berkualitas tinggi jika lebih banyak lebah yang mencari makan di area tersebut karena plot tersebut menyediakan habitat yang lebih baik sepanjang musim secara keseluruhan. Dengan demikian, kami akan melakukan jaring tangan yang ditargetkan untuk lebah di sebagian kecil lokasi pengambilan sampel pasif untuk digunakan dalam model kelimpahan. Jika ada korelasi positif antara jumlah lebah yang ditangkap oleh kedua metode, kami akan menggabungkan nilai-nilai ini untuk analisis (Bagian 2.8 ).
Kami akan menjaring lebah di tiga lokasi perangkap per pertanian musim panas setiap putaran pengambilan sampel (kecuali satu pertanian dengan hanya satu lokasi perangkap). Penangkapan lebah akan dilakukan pada siang hari dengan tutupan awan <50%, suhu >12 C dan angin rendah (Skala Angin Beaufort <4). Penangkapan lebah akan dilakukan oleh seorang pengamat tunggal yang berjalan pada empat garis transek 25-m yang sama yang digunakan untuk pengambilan sampel tanaman dan penghitungan pelet (Bagian 2.5 dan 2.6 ), mencari lebah 2 m di kedua sisi garis selama 30 menit (Gambar 3 ), dan menghentikan waktu pencarian untuk menangani atau memindahkan lebah ke stoples spesimen. Setelah survei, kami akan mencatat dan melepaskan setiap individu yang dapat diidentifikasi di lapangan, dan memindahkan mereka yang memerlukan sumber daya tambahan untuk identifikasi ke whirlpack dan identifikasi di laboratorium (Bagian 2.4 ).
2.4 Identifikasi lebah
Kami akan mengidentifikasi semua spesies lebah di lapangan atau laboratorium menggunakan Løken ( 1973 ), Løken ( 1985 ), Ødegaard et al. ( 2015 ), dan Rasmont et al. ( 2021 ). Karena identifikasi spesies untuk kompleks Bombus senso stricto bergantung pada analisis genetik (Murray et al., 2007 ; Waters et al., 2011 ), individu yang termasuk dalam kelompok ini tidak akan diidentifikasi hingga tingkat spesies. Semua spesimen serangga yang dibawa kembali ke laboratorium akan ditempatkan dalam freezer dan dipisahkan berdasarkan lokasi, metode penangkapan, dan hari pengambilan sampel hingga diproses. Setelah membilas, mengeringkan, dan menyematkan spesimen, kami akan mengidentifikasi semua lebah hingga spesies atau kompleks spesies di bawah pembesaran.
2.5 Pengambilan sampel tanaman dan tanah
Kami akan memperkirakan tutupan forb, tanah gundul, serasah, kelimpahan bunga dan tinggi rerumputan di area yang langsung mengelilingi setiap perangkap baling-baling biru pada empat transek 25 m yang memancar dari setiap perangkap selama periode pengambilan sampel masing-masing yang diidentifikasi di Bagian 2.2 (Gambar 3 ). Di titik tengah di bawah setiap perangkap, dan pada 10 dan 20 m di sepanjang setiap transek, kami akan memperkirakan tutupan forb, tanah gundul dan serasah sekali selama periode pramusim menggunakan kerangka sampel 50 x 50 cm dengan empat kabel internal yang bersilangan di setiap sisi. Jumlah kotak internal yang berisi setiap kategori akan dihitung saat dipegang setinggi pinggang, menghasilkan nilai total 0 hingga 25 untuk setiap kerangka dan dijumlahkan untuk setiap plot.
Untuk memperkirakan kelimpahan bunga selama setiap sesi pengambilan sampel, satu pengamat akan berjalan di setiap garis transek, mencari 1 m di kedua sisi, dan menghitung jumlah bunga yang menunjukkan struktur reproduksi matang (yaitu kepala sari, kepala putik, dan kelopak) untuk setiap spesies yang diserbuki oleh serangga (tumbuhan perdu dan semak). Untuk spesies dengan banyak bunga kecil, kami akan menghitung jumlah perbungaan. Kami akan menggabungkan hasil dari keempat transek, menghasilkan satu hitungan bunga per spesies tanaman per lokasi perangkap setiap putaran pengambilan sampel.
Terakhir, kami akan menilai tinggi rumput secara visual selama setiap sesi pengambilan sampel seperti yang dijelaskan dalam Milberg et al. ( 2016 ), mencatat tinggi vegetasi dalam tiga kategori (<5 cm, 5–15 cm dan >15 cm) per transek dan dinilai sebagai persentase dari semua empat transek 25-m yang digabungkan panjangnya. Seperti Milberg et al. ( 2016 ), kami juga akan menghitung metrik tinggi rumput rata-rata untuk setiap plot dengan terlebih dahulu mengubah kategori <5 cm menjadi 2,5 cm, 5–15 cm menjadi 10 cm dan >15 cm menjadi 25 cm, kemudian mengalikan nilai-nilai ini dengan persentase untuk setiap kategori dalam transek dan menjumlahkan hasilnya.
2.6 Metrik intensitas domba
Empat transek 25-m yang memancar dari setiap perangkap juga akan menjadi dasar hitungan pelet feses pengamat ganda selama masing-masing dari tiga sesi pengambilan sampel. Satu pengamat akan mencari 1 m di kedua sisi transek, mencatat dan membuang semua kelompok feses domba, diikuti oleh pengamat kedua yang menghitung dan membuang apa pun yang tidak ditemukan oleh pengamat pertama. Kami akan mempertimbangkan kelompok yang unik jika ada ≥3 pelet individu, feses yang menggumpal berjarak >20 cm dari satu sama lain atau jika warna, bentuk, dan tekstur menunjukkan usia pengendapan yang berbeda. Ini akan memberikan metrik intensitas domba jangka pendek untuk setiap plot. Untuk metrik intensitas domba jangka panjang kami, kami memperkirakan jumlah domba yang dilepaskan di setiap peternakan dengan mengalikan jumlah domba betina dengan jumlah rata-rata domba per jenis masing-masing seperti yang dilaporkan oleh petani, ditambah jumlah domba betina.
2.7 Klasifikasi metrik lanskap
Kami akan mendigitalkan total hektar habitat padang rumput terbuka di setiap pertanian musim panas secara manual dari foto udara di ArcGIS Pro 3.0.3 (ESRI, Redlands, CA, AS) untuk menghitung ukuran padang rumput. Kami juga akan mencatat koordinat spasial untuk setiap pusat plot dan menghitung jarak ke tepi hutan dari foto udara di GIS.
2.8 Analisis statistik
Secara umum, analisis kami akan meneliti penggembalaan domba sebagai pendorong kekayaan spesies lebah dan kelimpahan total. Kami akan melakukan semua analisis dalam versi R terkini (Tim Inti R, 2024 ) untuk penyerahan akhir pada Maret 2026. Semua data lapangan akan disertakan dalam analisis statistik kecuali inspeksi mengidentifikasi nilai ekstrem >3 SD. Kami akan menganggap data ini dicatat karena kesalahan dan mengecualikannya dari analisis lebih lanjut. Kami akan memeriksa variabel prediktor untuk multikolinearitas dengan menghitung Faktor Inflasi Varians dengan fungsi vif dalam paket mobil (Fox & Weisberg, 2019 ) dan mengecualikan yang bernilai >10 dari model yang sama (Hair et al., 2013 ).
Kami kemudian akan membangun serangkaian model Bayesian kandidat yang relevan secara ekologis menggunakan brm dalam paket brms (Bürkner, 2017 , 2018 , 2021 ) dengan kekayaan spesies lebah dan kelimpahan sebagai variabel respons kami dalam set model terpisah. Kami akan memeriksa distribusi data dengan histogram dan paket fitdistrplus (Delignette-Muller & Dutang, 2015 ) untuk memilih distribusi kesalahan yang tepat dan menggunakan default untuk rantai dan iterasi dengan prior informatif yang lemah untuk semua prediktor. Sebelum membangun model campuran kami, kami akan menghitung korelasi Pearson untuk menentukan kekuatan korelasi yang diantisipasi antara jaring tangan dan kelimpahan lebah yang berasal dari perangkap menggunakan fungsi cor.test dalam paket dasar R. Korelasi positif yang kuat ( r ≥ 0,7; Sokal & Rohlf, 1981 ) antara kelimpahan dari kedua metode penangkapan ini menunjukkan bahwa kedua metode tersebut sama-sama bias di seluruh pertanian musim panas. Dalam kasus ini, kami akan membangun model kelimpahan dari data jaring tangan saja dan juga kombinasi data dari perangkap dan jaring tangan. Jika tidak ditemukan korelasi, atau korelasi negatif, antara data dari kedua metode ini, ini akan menunjukkan perangkap baling-baling biru bukanlah metode yang dapat diandalkan untuk mengukur kelimpahan di lokasi kami dan model kelimpahan hanya akan dibangun menggunakan data dari jaring tangan. Jika kami mengamati perbedaan yang kuat dalam tingkat penangkapan, kami akan menghindari pola yang disebabkan oleh sejumlah kecil spesies dominan dengan membangun model kelimpahan khusus spesies untuk spesies yang paling umum.
Karena perkiraan jumlah domba yang dilepaskan di lokasi kami stabil dari waktu ke waktu, proporsi tumbuhan perdu, tanah kosong, dan serasah berubah dalam skala multi-tahun, dan jumlah tanah kosong dan serasah relevan dengan ratu lebah hanya selama periode perkembangan sarang, kami akan menyertakan prediktor yang diukur hanya sekali dalam model yang dirancang untuk menguji efek jangka panjang penggembalaan domba. Sebaliknya, jumlah domba yang hadir pada skala plot selama periode pengambilan sampel tertentu dalam musim yang dijelaskan oleh metrik jumlah pelet feses kami, jumlah dan kekayaan spesies bunga, dan tinggi rerumputan semuanya merupakan ukuran yang berhubungan dengan efek jangka pendek langsung dari penggembalaan domba. Dengan demikian, kami akan mengidentifikasi model yang bersaing untuk kekayaan dan kelimpahan spesies lebah yang memungkinkan perbandingan pada salah satu dan kedua skala ini. Untuk menguji hipotesis kami bahwa pengaruh penggembalaan domba terhadap sumber daya mencari makan merupakan penentu yang lebih kuat dari kekayaan dan kelimpahan spesies lebah dalam sistem kami daripada karakteristik yang mendorong ketersediaan lokasi sarang, kami juga akan membandingkan model dengan variabel mencari makan saja dengan yang mencakup variabel mencari makan dan lokasi sarang. Kami akan mempertimbangkan berbagai intersepsi pada sesi pengambilan sampel, plot individual dan pertanian musim panas serta interaksi relevan dan efek non-linier (kuadrat). Model dan prediksi individual dirinci dalam Tabel 2. Untuk mengidentifikasi yang terbaik di antara model yang bersaing, kami akan memilih model yang memiliki nilai kriteria informasi validasi silang (looic) leave-one-out terendah dengan fungsi looic dan loo_compare dalam paket loo (Vehtari et al., 2016 , 2024 ). Kami akan memeriksa plot jejak dan R-hat untuk menilai konvergensi model dan memeriksa distribusi prediktif posterior untuk menilai kebaikan kecocokan dalam paket shiny_stan (Gabry & Veen, 2022 ). Kami akan melaporkan plot koefisien dan prediksi untuk model yang kami pilih.
| Hipotesis yang menjelaskan kekayaan dan kelimpahan spesies lebah ( y ) | Komponen model a |
|---|---|
| Jumlah domba (yang dilepas) pada skala peternakan musim panas menjelaskan y karena tekanan penggembalaan jangka panjang | Jumlah domba |
| Jumlah pelet domba per putaran pengambilan sampel, sebagai indikator intensitas penggembalaan jangka pendek (dan langsung), menjelaskan y | Jumlah pelet |
| Variabel jangka panjang yang hanya berhubungan dengan sumber daya mencari makan dan ditentukan oleh jumlah domba menjelaskan y | Jumlah domba + Luas padang rumput + Penutupan forb |
| Variabel jangka panjang yang berhubungan dengan sumber daya mencari makan dan bersarang yang ditentukan oleh jumlah domba menjelaskan mengapa | Jumlah domba + Luas padang rumput + Penutupan forb + Serasah + Tanah gundul + Jarak ke hutan |
| Variabel jangka pendek yang hanya berhubungan dengan sumber daya mencari makan dipengaruhi oleh intensitas penggembalaan domba per putaran pengambilan sampel menjelaskan y | Jumlah pelet * Kekayaan spesies bunga + Jumlah bunga * Tinggi rerumputan |
| Kombinasi variabel jangka pendek dan jangka panjang yang hanya berhubungan dengan sumber daya mencari makan dan dipengaruhi oleh jumlah domba pada skala peternakan musim panas, menjelaskan y | Jumlah domba * Jumlah bunga + Penutupan forb + Kekayaan spesies bunga + Jumlah bunga * Tinggi rerumputan + Luas padang rumput + Penutupan forb |
| Kombinasi variabel jangka pendek dan jangka panjang yang hanya berkaitan dengan sumber daya mencari makan dan dipengaruhi oleh intensitas penggembalaan domba per putaran pengambilan sampel menjelaskan y | Jumlah pelet * Penutupan forb + Kekayaan spesies bunga + Jumlah bunga * Tinggi rerumputan + Ukuran padang rumput + Penutupan forb |
| Semua variabel habitat yang diukur pada skala plot dan lanskap yang penting untuk mencari makan dan bersarang, dipengaruhi oleh jumlah domba pada skala pertanian musim panas, menjelaskan mengapa | Jumlah domba * Penutup padang rumput + Jumlah domba * Serasah + Tanah kosong + Jumlah bunga * Tinggi padang rumput + Kekayaan spesies bunga + Jarak ke hutan + Luas padang rumput |
| Semua variabel habitat yang diukur pada skala plot dan lanskap yang penting untuk mencari makan dan bersarang, dipengaruhi oleh intensitas domba per putaran pengambilan sampel, menjelaskan mengapa | Jumlah pelet * Jumlah bunga + Penutupan forb + Kekayaan spesies bunga + Jumlah bunga * Tinggi rerumputan + Serasah + Tanah kosong + Jarak ke hutan + Ukuran padang rumput |
| Semua variabel habitat yang diukur pada skala plot dan lanskap yang penting untuk mencari makan dan bersarang, terlepas dari domba, menjelaskan mengapa | Penutupan rumput + Serasah + Tanah kosong + Kekayaan spesies bunga + Jumlah bunga * Tinggi padang rumput + Jarak ke hutan + Luas padang rumput |
| Model nol | Hanya istilah intersepsi acak |
Catatan : Semua model mempertimbangkan intersepsi yang bervariasi pada sesi pengambilan sampel, plot individu, dan peternakan musim panas. Variabel prediktor yang diambil sampelnya sekali sebelum pelepasan domba, sehingga memiliki satu nilai untuk musim tersebut dan berlaku dalam jangka panjang, dicatat dalam huruf miring. Variabel prediktor yang dimodelkan sebagai kuadrat dicatat dalam huruf tebal. a Semua model berisi istilah intersepsi acak untuk pertanian musim panas, transek, dan putaran pengambilan sampel.
3 DISKUSI
Meskipun padang rumput semi-alami diakui sebagai habitat penting bagi lebah di Fennoscandia, hingga kini belum jelas bagaimana mereka menanggapi berbagai tingkat gangguan oleh domba di lanskap boreal konifer. Hasil penyelidikan kami akan memberikan informasi berharga kepada pengelola lahan mengenai jumlah domba liar yang akan dilepaskan di padang rumput semi-alami di lanskap boreal dan juga akan berkontribusi pada pemahaman yang lebih baik tentang penggembalaan ternak sebagai alat konservasi secara umum.