
Abstrak
- Perambahan semak belukar menimbulkan tantangan yang signifikan di banyak ekosistem padang rumput, terutama di tempat-tempat yang sering terjadi gangguan seperti penggembalaan ternak yang besar dan kebakaran yang tidak terkendali. Kebakaran sering digunakan sebagai alat manajemen untuk mengurangi perambahan semak belukar dan melestarikan vegetasi asli, karena kebakaran memengaruhi pertumbuhan kembali dan perkecambahan biji. Namun, hanya sedikit penelitian yang meneliti dampak waktu kebakaran terhadap pertumbuhan kembali dan perkecambahan biji baik di lapangan maupun di tempat kejadian.
- Kami menyelidiki kemampuan perkecambahan semak Helichrysum setelah terbakar seluruhnya, sebagian atau tidak terbakar sama sekali. Selain itu, kami mengumpulkan sampel bank benih tanah dari setiap area yang terbakar awal, terbakar akhir dan tidak terbakar pada kedalaman 0–2, 2–4 dan 4–6 cm untuk menilai perekrutan spesies tanaman melalui pembibitan. Data dianalisis menggunakan model regresi linier umum dengan fungsi tautan Gaussian.
- Kami menemukan perbedaan yang signifikan dalam jumlah tunas baru dan kelimpahan bibit di lokasi yang terbakar awal, terbakar akhir, dan tidak terbakar. Tunas baru dari semak yang terbakar sebagian dan terbakar total lebih banyak dibandingkan dengan semak yang tidak terbakar. Tanah permukaan di lokasi yang tidak terbakar mengandung lebih banyak bibit Helichrysum dibandingkan dengan lokasi yang terbakar, yang menunjukkan bahwa kebakaran berdampak negatif pada perkecambahan benih, terutama di lapisan tanah atas. Secara keseluruhan, perekrutan bibit lebih rendah di area yang terbakar awal dan akhir, sedangkan tunas baru lebih tinggi di area yang terbakar di akhir musim.
- Implikasi praktis . Temuan ini menyoroti pentingnya pengaturan waktu kebakaran dalam pengelolaan padang rumput. Kami menganjurkan penerapan pembakaran dini yang terencana untuk mengurangi perambahan semak belukar, meningkatkan keanekaragaman hayati, dan menyelamatkan padang rumput yang sensitif secara ekologis.
1. PENDAHULUAN
Perambahan semak suksesi sekunder merupakan masalah yang umum terjadi di sebagian besar ekosistem padang rumput yang gangguan sebelumnya telah berhenti (Stankeva Terziyska et al., 2020 ). Penggerak utama perambahan semak adalah perubahan dalam kebakaran dan rezim penggembalaan (Mapiye et al., 2008 ). Perluasan semak ke padang rumput menggusur spesies herba asli karena persaingan untuk mendapatkan cahaya, kelembaban, dan nutrisi (Manish et al., 2016 ; Ruprecht et al., 2015 ). Perambahan ini menimbulkan masalah konservasi ekologi yang signifikan (Pornaro et al., 2017 ), khususnya di kawasan lindung dan wilayah dengan keanekaragaman spesies tanaman dan endemisme yang tinggi (Guido et al., 2017 ).
Di Afrika timur, api telah digunakan selama beberapa dekade sebagai alat manajemen untuk melestarikan ekosistem padang rumput dengan mengendalikan perambahan semak belukar (Nyakunga et al., 2018 ). Api yang direncanakan (pembakaran awal) biasanya dilakukan pada awal musim kemarau ketika beberapa rumput dan herba masih hijau dan tanah mempertahankan kelembaban yang memadai. Kondisi ini menghasilkan api yang relatif dingin (Bloesch, 1999 ). Sebaliknya, kebakaran hutan bersuhu tinggi, yang sering kali tidak terkendali, biasanya terjadi pada akhir musim kemarau ketika tingkat kelembaban tanaman rendah dan fenomena ini disebut sebagai ‘pembakaran akhir’ (Bloesch, 1999 ; Govender et al., 2006 ; Tangney et al., 2022 ).
Spesies tanaman yang berbeda bervariasi dalam respons mereka saat terpapar pada rezim kebakaran yang berbeda (Drewa et al., 2002 ). Di lingkungan yang rawan kebakaran, tanaman bertahan hidup melalui perekrutan dari bank benih tanah atau melalui resprouting (resprouter obligat pasca-kebakaran) untuk mempertahankan populasi mereka (Luo et al., 2022 ). Resprouter obligat bergantung pada resprouting untuk bertahan hidup setelah peristiwa kebakaran menghancurkan benih mereka (Marais et al., 2014 ). Sebaliknya, seeder obligat binasa setelah kebakaran tetapi dapat beregenerasi secara eksklusif dari benih yang disimpan di dalam tanah (Prior & Bowman, 2020 ), menjadikan regenerasi benih satu-satunya cara mereka untuk bertahan hidup (Tangney et al., 2022 ). Seeder fakultatif dan resprouter fakultatif dapat merekrut dari benih dan bertunas kembali setelah kebakaran (Luo et al., 2022 ). Beberapa resprouter obligat pasca-kebakaran berhasil bertahan hidup dari kebakaran dan beregenerasi dari bank tunas di atas atau di bawah tanah (Clarke et al., 2013 ), menjadikan regenerasi benih sebagai satu-satunya cara mereka untuk bertahan hidup (Tangney et al., 2022 ). Struktur di bawah tanah seperti lignotuber dan akar memainkan peran penting dalam menyimpan karbohidrat, gula, dan nutrisi non-struktural, yang memfasilitasi resprouting setelah kebakaran (Clarke et al., 2013 ). Namun, kapasitas untuk resprouting setelah kebakaran juga dipengaruhi oleh frekuensi dan tingkat keparahan kebakaran (Martínková et al., 2004 ). Ketika bagian tanaman di atas tanah rusak atau dihilangkan, tunas dorman atau jaringan meristematik yang terletak di organ di bawah tanah diaktifkan untuk menghasilkan tunas dan daun baru (Thomsen & Ooi, 2022 ). Faktor-faktor yang berkontribusi terhadap respons resprouting ini termasuk aktivasi tunas dan ketersediaan sumber daya yang tersimpan. Spesies yang tumbuh kembali menunjukkan ketahanan yang lebih besar terhadap kebakaran panas dibandingkan dengan spesies yang tumbuh kembali (Bennett et al., 2016 ). Waktu kebakaran memengaruhi kekuatan tanaman untuk tumbuh kembali (Thomsen & Ooi, 2022 ), dengan suhu kebakaran yang tinggi mengganggu perkecambahan benih sementara suhu kebakaran yang rendah umumnya mendorong pertumbuhan kembali.
Pengelolaan kebakaran digunakan di banyak Taman Nasional untuk menekan masuknya spesies semak atau pohon berkayu (Brancatelli et al., 2024 ; Ling et al., 2023 ). Masuknya spesies semak ini juga telah diamati di Taman Nasional Kitulo (KNP) di Tanzania (URT, 2010 ), sebuah taman yang terkenal akan keanekaragaman floranya yang tinggi dan rumah bagi lebih dari 85 spesies anggrek darat yang hidup di padang rumput (Davenport & Markes, 2018 ; Davenport & Ndangalasi, 2003 ; Veldman et al., 2018 ). Perambahan ini merupakan ancaman potensial bagi anggrek ini (Ndaki et al., 2021 ), karena sebagian besar spesies lebih menyukai habitat padang rumput terbuka dan dapat terpengaruh secara negatif oleh perambahan tersebut (Martín-Forés et al., 2022 ; Slaviero et al., 2016 ; Vogt-Schilb et al., 2015 ). Kebakaran yang direncanakan dan kebakaran hutan telah terjadi di TNK selama lebih dari satu dekade dengan tujuan untuk mencegah kebakaran hutan yang merusak (Mwinuka & Mushi, komunikasi pribadi) tetapi waktu kebakaran agak acak atau diabaikan dalam pendekatan manajemen. Namun, karena waktu kebakaran sering kali menentukan panas api tetapi juga memengaruhi tanaman dalam tahap fenologi yang berbeda (Richardson & Wagenius, 2022 ), waktu mungkin memiliki konsekuensi yang cukup parah pada spesies tanaman mana yang dapat bertahan hidup dari peristiwa kebakaran. Dalam studi ini, ‘pembakaran akhir’ mengacu pada kebakaran hutan dan ‘pembakaran awal’ menunjukkan kebakaran yang direncanakan. Pembakaran akhir biasanya terjadi pada musim yang lebih hangat atau kering ketika bahan organik kering, sehingga menghasilkan kebakaran berintensitas tinggi pada suhu panas (Mackenzie et al., 2021 ). Sebaliknya, pembakaran awal terjadi pada awal musim kemarau ketika beberapa rumput dan herba masih hijau dan tanah masih memiliki kelembapan yang cukup.
Salah satu spesies yang merambah yang dilaporkan di TNK adalah Helichrysum splendidum , yang paling dominan dibandingkan spesies Helichrysum lainnya (Mgimba et al., 2024 ). Spesies ini dapat diperbanyak baik melalui biji maupun stek (Giovannini et al., 2008 ), sementara spesies Helichrysum lainnya dapat tumbuh kembali dari akar (Makena et al., 2023 ). Helichrysum foetidum (L.) Moench, Helichrysum aureonitens Sch.Bip. dan Helichrysum vestitum beregenerasi melalui tunas kembali dan bersifat ephemeral api dan sklerofil, dengan perkecambahan biji difasilitasi oleh api (Afolayan et al., 1997 ; Brown, 1992 ; Riveiro et al., 2019 ). Penelitian telah menunjukkan bahwa benih Helichrysum cassinianum dan Helichrysum odoratissimum berkecambah secara optimal pada suhu udara antara 15 dan 20°C (Makena et al., 2023 ; Mott, 1974 ). Di sisi lain, penelitian tentang pengaruh kebakaran terhadap strategi persistensi Helichrysum trilineatum menunjukkan bahwa spesies ini memiliki vigor resprouting yang tinggi setelah kebakaran (Kraaij et al., 2017 ). Hingga saat ini, informasi tentang potensi resprouting belum cukup dieksplorasi pada Helichrysum splendidum , yang dominan di wilayah penelitian. Lebih jauh, meskipun beberapa penelitian telah menyelidiki efek suhu kebakaran pada perkecambahan benih berbagai spesies Helichrysum (Afolayan et al., 1997 ; Giovannini et al., 2008 ; Makena et al., 2023 ; Mott, 1972 ; Riveiro et al., 2019 ), hanya sedikit yang mengeksplorasi efek terjadinya kebakaran pada bank benih Helichrysum di dalam tanah (Wube et al., 2021 ). Kami berhipotesis bahwa tingkat hangusnya batang Helichrysum akibat kebakaran memiliki efek pada regenerasinya melalui resprouting. Kami juga berhipotesis bahwa pembakaran yang terlambat menghambat regenerasi spesies Helichrysum melalui perekrutan bibit. Untuk menguji hipotesis ini, kami melakukan studi observasional pada semak Helichrysum di bawah terjadinya kebakaran di KNP, Tanzania. Temuan kami akan membantu kami memahami bagaimana manajemen kebakaran memengaruhi perambahan Helichrysum spp. di sistem padang rumput dan memberikan saran tentang waktu kejadian kebakaran.
2 BAHAN DAN METODE
2.1 Studi deskripsi spesies tanaman
Spesies yang teridentifikasi di TNK adalah Helichrysum splendidum , Helichrysum odoratissimum , Helichrysum forskahlii dan Helichrysum kirkii (Mgimba et al., 2024 ; Tabel 1 ).
| Jenis | Bentuk kehidupan | Regenerasi | Anatomi | Referensi |
|---|---|---|---|---|
| H. forskahlii | Herba atau semak abadi | Reprouter fakultatif | Batang berkayu, banyak cabang | Beentje ( 2002 ), Makena dkk. ( 2023 ) |
| H. odoratissimum | Ramuan abadi | Reprouter fakultatif | Batang berkayu, tegak atau menyebar, seringkali memiliki banyak cabang | Serabele dkk. ( 2023 ), Makena dkk. ( 2023 ) |
| H. kirkii | Ramuan abadi | Reprouter fakultatif | Beberapa batang tegak, dengan cabang tunggal atau sedikit, penutup daun rapat | Beentje ( 2000 ) |
| Bunga H. yang indah | Semak abadi | Reprouter fakultatif | Tinggi, dengan cabang-cabang yang menghasilkan daun-daun berbulu abu-abu tipis | Roux ( 2003 ), Steger dan kawan-kawan ( 2022 ) |
Spesies Helichrysum menghasilkan benih ringan yang mudah disebarkan oleh angin (Chengere et al., 2023 ). Meskipun sebagian besar bank benih terletak di permukaan tanah, kepadatan dan viabilitas benih menurun seiring bertambahnya kedalaman tanah (Wakshum et al., 2018 ). Distribusi vertikal bank benih tanah dipengaruhi oleh berbagai faktor, termasuk jenis tanah, ukuran benih, dan bentuk benih (Csontos, 2007 ). Misalnya, benih yang lebih besar cenderung tidak menembus lapisan tanah yang lebih dalam melalui pergerakan di sepanjang rekahan atau penguburan oleh hewan penghuni tanah, dibandingkan dengan benih yang lebih kecil (Birhanu et al., 2022 ). Selain itu, spesies tanaman dengan benih yang menunjukkan umur lebih panjang dapat bertahan di lapisan tanah yang lebih dalam (Wakshum et al., 2018 ). Kebakaran juga memiliki efek panas yang lebih besar di tanah yang lebih dekat ke permukaan daripada di tanah yang lebih dalam (Badía et al., 2017 ) karena tanah merupakan konduktor panas yang buruk (DeBano, 2000 ). Memahami dampak kebakaran pada spesies Helichrysum sangat penting untuk pengelolaan ekologi, khususnya di padang rumput Taman Nasional Tanzania, tempat semak Helichrysum berkembang biak.
2.2 Deskripsi wilayah studi
Penelitian ini dilakukan di TNK, yang terletak di dataran tinggi Selatan Tanzania (09°09′15″ S dan 33°57′05″ E; Gambar 1 ). Taman ini meliputi dataran tinggi pada ketinggian 2500 m dpl (Lovett & Prins, 1994 ; URT, 2010 ) dan dicirikan oleh padang rumput pegunungan (Nyomora, 2009 ). Periode musim panas (November–April) adalah musim hujan, dengan curah hujan tahunan rata-rata berkisar antara 1500 hingga 2900 mm dan suhu rata-rata sekitar 18°C (Davenport & Markes, 2018 ). Musim dingin (Mei–Oktober) kering, dengan suhu rata-rata sekitar 15°C (URT, 2010 ). Fluktuasi suhu yang signifikan terjadi selama musim dingin, dengan suhu malam hari terkadang turun di bawah 0°C (URT, 2020 ).
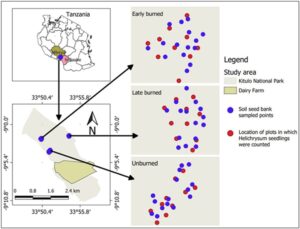
Sebelum tahun 1965, padang rumput dalam ekosistem TNK dimanfaatkan oleh masyarakat lokal untuk penggembalaan ternak dan budidaya tanaman seperti Tanacetum cinerariifolium (piretrum), Solanum tuberosum L. (kentang Irlandia) dan Triticum aestivum L. (gandum) (Lovett & Prins, 1994 ). Produksi serealia komersial dan domba dihentikan pada tahun 1968 dan 1973, masing-masing (Lovett & Prins, 1994 ). Selanjutnya, hanya produksi susu komersial dan tanaman skala kecil yang berlanjut hingga deklarasi TNK sebagai taman nasional pada tahun 2005 (Mwinuka dan Mushi, komunikasi pribadi).
Kebakaran dini terjadi di TNK kadang-kadang pada awal Juni, seperti yang ditentukan oleh manajemen dan kebakaran hutan terjadi kadang-kadang sepanjang tahun tetapi terutama antara akhir Agustus hingga Oktober. Kebakaran dini terjadi ketika vegetasi masih hijau; dengan demikian, peristiwa kebakaran ini memiliki suhu dan intensitas yang agak rendah karena fakta bahwa sebagian energi yang dilepaskan digunakan untuk menguapkan kelembaban yang tersisa dari vegetasi (Bloesch, 1999 ). Sebaliknya, peristiwa kebakaran akhir memiliki suhu dan intensitas yang tinggi karena bahan organik kering yang menambah beban bahan bakar (Bloesch, 1999 ; Govender et al., 2006 ).
2.3 Dampak kebakaran terhadap perkecambahan individu Helichrysum
Bahasa Indonesia: Untuk menyelidiki dampak kebakaran pada perkecambahan Helichrysum spp., tiga plot berukuran 50 × 50 m, yang diberi jarak 300 m, ditandai secara permanen di dalam area yang didominasi oleh Helichrysum spp. Lokasi ini telah mengalami kebakaran akhir selama dua tahun berturut-turut, yang berarti mengalami kebakaran panas karena vegetasi kering mudah terbakar. Lokasi tersebut terbakar akhir lagi pada bulan September 2021, dan penilaian perkecambahan dimulai 2 bulan kemudian, pada bulan November 2021. Di dalam setiap plot, setiap individu Helichrysum yang dapat kami identifikasi dikategorikan berdasarkan tingkat kerusakan akibat kebakaran (Morrison, 2002 ; Vesk & Westoby, 2004 ). Individu Helichrysum didefinisikan sebagai rimpang tunggal, yang darinya dapat muncul beberapa batang (Pljevljakušić et al., 2018 ). Kami tidak dapat mengidentifikasi semua spesies Helichrysum hingga tingkat spesies dalam penilaian dampak kebakaran karena empat spesies utama yang telah kami identifikasi dalam studi sebelumnya (Mgimba et al., 2024 ) sering ditemukan hidup berdampingan. Fenologi mereka yang serupa pada tahap pembibitan juga membuat identifikasi menjadi sulit. Kategori yang kami nilai termasuk terbakar sebagian, meliputi individu dengan batang hangus kurang dari 50%; terbakar sepenuhnya, meliputi individu dengan batang hangus lebih dari 0% dan tidak terbakar, terdiri dari individu yang tidak hangus. Dalam setiap kategori, 10 individu per plot diambil sampelnya ( N = 90). Untuk setiap individu yang diambil sampelnya, setiap tunas yang bertahan hidup ditandai dengan selotip, dan jumlah tunas kembali dicatat setiap bulan selama periode 5 bulan, dari November 2021 hingga Maret 2022.
2.4 Dampak kebakaran terhadap perekrutan bibit Helichrysum
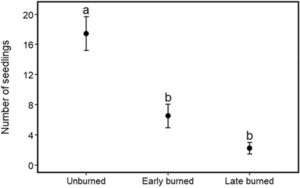
Untuk mengevaluasi dampak waktu kebakaran pada benih spesies Helichrysum , kami menetapkan lokasi yang tidak terbakar selama lebih dari dua tahun berturut-turut sebagaimana diamati dari data bekas kebakaran yang diindera dari jarak jauh yang diunduh ( http://earthexplorer.usgs.gov ). Di dalam area ini, kami memilih dua lokasi masing-masing seluas 6,7 ha (satu untuk kebakaran awal dan lainnya untuk kebakaran akhir), dengan mempertimbangkan bahwa lokasi-lokasi ini telah mengalami kejadian kebakaran yang serupa selama dua tahun berturut-turut. Di setiap lokasi, sepuluh kuadrat berukuran 5 m × 5 m yang didominasi oleh Helichrysum spp. ditetapkan secara acak pada bulan Februari 2022 pada jarak 250 m dari satu sama lain, N = 30 (Guido et al., 2017 ). Jumlah bibit Helichrysum kemudian dicatat di setiap kuadrat. Bibit Helichrysum didefinisikan sebagai tanaman muda <2 cm tinggi (Harris et al., 2022 ). 2.5 Dampak kebakaran terhadap persediaan benih tanah spesies Helichrysum Bahasa Indonesia: Untuk memahami bagaimana waktu kejadian kebakaran mempengaruhi bank benih tanah, kami mengumpulkan sampel bank benih tanah dari area yang terbakar awal, terbakar akhir, dan tidak terbakar (Gambar 1 ). Area yang terbakar awal dikelola oleh Otoritas Taman Nasional Tanzania TANAPA, dan area yang terbakar akhir disebabkan oleh kebakaran hutan yang tidak disengaja. Di setiap area, kami membuat lima belas kuadrat berukuran 5 m × 5 m di wilayah dengan individu Helichrysum yang melimpah (Guido et al., 2017 ). Kami mengumpulkan sampel tanah pada tiga kedalaman: 0–2 cm (tanah permukaan), 2–4 dan 4–6 cm (Bekele et al., 2022 ; Savadogo et al., 2017 ), diambil dari bagian tengah dan sudut setiap kuadrat, lalu digabungkan untuk membentuk sampel tanah komposit sekitar 1000 g ( N = 45). Sampel tanah dikeringkan pada suhu 24–27°C selama 2 minggu dan diayak melalui saringan 4 mm untuk membuang serpihan (Wube et al., 2021 ). Tanah yang diayak dimasukkan ke dalam wadah plastik berlubang yang diletakkan di ruang terbuka dan disiram setiap 2 hari (Wube et al., 2021 ). Selama periode 2 bulan, dari Juli 2022 hingga September 2022, kami memantau dan mengidentifikasi kemunculan bibit tanaman setiap minggu (Harris et al., 2021 ). Bibit yang tercatat dibuang untuk menghindari penghitungan ulang. Kelimpahan bank benih tanah Helichrysum diperkirakan berdasarkan jumlah kumulatif bibit yang muncul selama seluruh periode pengamatan. 2.6 Analisis statistik Mengingat beberapa pengukuran dilakukan pada individu yang sama, kami menyertakan pengenal unik (ID; N = 10) untuk setiap tunas kembali. ID tunas kembali disarangkan dalam plot ( N = 3) untuk memperhitungkan perbedaan potensial antara plot eksperimen. Kami menggunakan model campuran linear umum dengan binomial negatif (Fahrmeir & Kneib, 2011 ; Gałecki & Burzykowski, 2013 ) untuk membandingkan jumlah total tunas kembali di antara semak yang tidak terbakar, terbakar sebagian, dan terbakar total pada akhir eksperimen (Maret 2022). Karena peraturan yang ketat di kawasan lindung, penanganan kebakaran tidak dapat direplikasi, sehingga menimbulkan potensi masalah replikasi semu (Hurlbert, 1984 ). Meskipun kriteria informasi Akaike (AIC) untuk model distribusi Poisson sedikit lebih rendah daripada AIC untuk dispersi binomial negatif, untuk kedua model itu >1 (yaitu 2,6). Kami berasumsi bahwa model binomial negatif lebih tangguh daripada model Poisson karena model sebelumnya memperhitungkan penyebaran berlebih (Hilbe, 2011 ). Kami mengendalikan variasi individual dan spasial dengan memasukkan ID tunas kembali yang bersarang di dalam plot sebagai efek acak (Fahrmeir & Kneib, 2011 ). Kami juga mengilustrasikan tren tunas kembali di bawah berbagai tingkat kehangusan dari waktu ke waktu sebagai kemungkinan interaksi antara kebakaran dan bulan dalam model, untuk menyelidiki apakah semak yang terpengaruh oleh berbagai perlakuan kebakaran menghasilkan tunas kembali baru secara berbeda dari waktu ke waktu. Kami mengendalikan variasi individual dan spasial dengan memasukkan ID tunas kembali yang bersarang di dalam plot sebagai efek acak. Untuk membandingkan jumlah benih Helichrysum di bank benih tanah di seluruh perlakuan kebakaran dan kedalaman tanah, kami menggunakan model regresi efek campuran linier umum dengan fungsi binomial negatif (Faraway, 2016 ) untuk memperhitungkan penyebaran berlebih, menggabungkan interaksi antara perlakuan kebakaran dan lapisan tanah. Waktu kebakaran, kedalaman tanah dan interaksi ketiganya dimasukkan sebagai efek tetap, sementara plot dimasukkan sebagai faktor acak untuk mengendalikan potensi variasi spasial antara plot.
Terakhir, kami menggunakan model regresi efek campuran linier umum dengan fungsi tautan Poisson untuk membandingkan kelimpahan bibit (variabel jumlah) di seluruh lokasi yang terpapar waktu kebakaran yang berbeda. Untuk memperhitungkan potensi variasi dalam plot dan subplot eksperimen, kami menyertakan subplot yang bersarang di dalam plot sebagai efek acak. Kami menggunakan R (versi 4.1.3) dan paket vegan (Sengupta, 2001 ) untuk analisis data dan perangkat lunak Origin Pro versi 2019 (9.65) (Speight, 2005 ) untuk membuat grafik.
3 HASIL
3.1 Regenerasi Helichrysum melalui perkecambahan setelah kebakaran
Model efek campuran menunjukkan bahwa pertumbuhan kembali pada batang Helichrysum yang terbakar berbeda secara signifikan. Individu Helichrysum yang sebagian atau bahkan seluruhnya terbakar oleh api menunjukkan kapasitas regeneratif hampir dua kali lipat melalui pertumbuhan kembali dibandingkan individu yang tidak terbakar (Tabel 2 ; Gambar 2A ).
| Memperkirakan | Bahasa Inggris | dari | P | |
|---|---|---|---|---|
| (Mencegat) | 0,475 tahun | 0,064 tahun | 7.375 | <0,001 |
| Kerusakan total | 0,255 | 0,086 tahun | 2.969 | 0,003 |
| Kerusakan sebagian | 0.222 | 0,086 tahun | 2.565 | 0,010 * |
* p < 0,05.

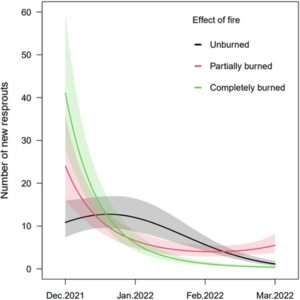
Dua bulan pasca kebakaran, jumlah tunas baru di semak yang tidak terbakar kira-kira setengah dari yang terbakar total dan sebagian ( t = 13,0, df = 58 p < 0,001; t = 10,7, p < 0,001, df = 58; Gambar 2A ). Proporsi ini tetap sama hingga Maret, yaitu, 5 bulan pasca kebakaran, dengan jumlah tunas baru di semak yang tidak terbakar kira-kira tiga kali lebih rendah daripada di semak yang terbakar total ( t = −2,4, p = 0,05, df = 58; Gambar 2B ). Pada bulan Desember, yaitu, 3 bulan setelah kebakaran, jumlah rata-rata tunas baru dari semak yang terbakar total kira-kira empat kali lebih tinggi daripada di semak yang tidak terbakar ( z = 5,0, p < 0,001). Namun, jumlah tunas baru menurun drastis dari waktu ke waktu di semua perlakuan (Gambar 3 ).
3.2 Kelimpahan bank benih tanah Helichrysum
Jumlah benih Helichrysum di tanah pada lokasi yang tidak terbakar dua kali lebih tinggi dibanding lokasi yang terbakar pada awal kebakaran dan 10 kali lebih tinggi dibanding lapisan tanah permukaan pada lokasi yang terbakar pada akhir kebakaran (Tabel 3 ; Gambar 4 ).
| Memperkirakan | Bahasa Inggris | dari | P | |
|---|---|---|---|---|
| Mencegat | 1.828 | 0.120 | 15.184 | <0,001 |
| Terbakar terlambat | -1,083 | 0.142 | -7.626 | <0,001 |
| Tidak terbakar | 0,987 | 0,084 tahun | 11.787 | <0,001 |
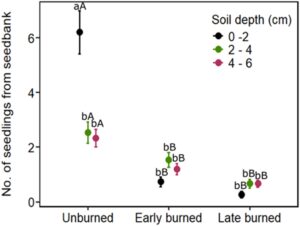
Inkubasi bank benih tanah kami memperlihatkan bahwa jumlah benih di tanah yang telah mengalami pembakaran awal dan akhir hanya muncul dalam setengah kasus dibandingkan dengan yang ada di tanah yang tidak terbakar. Efek ini paling kuat terlihat untuk benih di lapisan tanah permukaan dibandingkan dengan lapisan tanah yang lebih dalam (Tabel 4 ). Di lokasi yang terbakar awal dan akhir, kelimpahan bank benih per plot (25 m 2 ) tidak berbeda secara signifikan di seluruh kedalaman tanah, kecuali untuk lokasi yang tidak terbakar (Gambar 5 ). Di lokasi yang tidak terbakar, kelimpahan bank benih di tanah permukaan hampir tiga kali lebih tinggi daripada di kedalaman tanah yang lebih rendah, dengan lapisan 2–4 dan 4–6 cm memiliki kelimpahan bank benih yang hampir sama (Gambar 5 ).
| Memperkirakan | Bahasa Inggris | dari | P | |
|---|---|---|---|---|
| (Mencegat) | -0,065 | 0,258 | -0,250 | 0.803 |
| Terbakar lambat (0–2 cm) | -1.609 | 0.632 | -2.545 | 0,011 * |
| Tidak terbakar (0–2 cm) | 2.425 | 0.270 | 8.979 | <0,001 |
| Terbakar dini (2–4 cm) | 0,065 tahun | 0,365 tahun | 0,177 tahun | 0.860 |
| Terbakar lambat (2–4 cm) | -0,852 | 0.483 | -1.763 | 0,078 tahun |
| Tidak terbakar (2–4 cm) | 0.615 | 0.324 | 1.895 | 0,058 |
| Terbakar dini (4–6 cm) | -1,034 | 0.516 | -2,002 | 0,045 * |
| Terbakar lambat (4–6 cm) | -1.545 | 0.632 | -2.443 | 0,015 * |
| Tidak terbakar (4–6 cm) | -0,446 | 0.422 | -1,058 | 0.299 |
* p < 0,05.
4 DISKUSI
4.1 Pengaruh kebakaran terhadap kemampuan perkecambahan spesies Helichrysum
Bertentangan dengan harapan kami, kami menemukan bahwa semak Helichrysum yang terbakar seluruhnya menunjukkan jumlah tunas ulang yang lebih banyak dibandingkan dengan semak yang terbakar sebagian dan tidak terbakar. Kami berharap bahwa tanaman Helichrysum yang tidak terbakar akan menunjukkan kekuatan tunas ulang yang lebih besar daripada yang terbakar dan bahwa hangus total akan menghasilkan sedikit atau tidak ada tunas ulang karena kebakaran sangat menekan tunas ulang pada spesies tanaman lain, misalnya, Manilkara sansibarensis dan Baphia kirkii (Mligo, 2019 ). Mirip dengan temuan kami, Strydom et al. ( 2020 ) menilai kelangsungan hidup dan kekuatan tunas ulang pasca-kebakaran dalam kaitannya dengan tingkat keparahan kebakaran dan menemukan bahwa tingkat keparahan kebakaran memiliki efek positif pada jumlah tunas ulang semak belukar di Afrika Selatan. Lebih lanjut, Schafer dan Just ( 2014 ) menemukan bahwa 95% tanaman semak tumbuh kembali setelah terbakar seluruhnya dan pembuangan biomassa di atas tanah dalam sebuah penelitian di Carolina Utara. Demikian pula, spesies tanaman berkayu di lokasi yang terbakar dan tidak terbakar menunjukkan kemampuan tumbuh kembali yang lebih tinggi di lokasi yang terbakar dibandingkan di area Mediterania yang tidak terbakar di Spanyol Timur Laut (Quevedo et al., 2007 ). Penghapusan biomassa di atas tanah mengalokasikan kembali karbohidrat yang tersimpan di akar tanaman untuk produksi dan pertumbuhan batang baru (Clarke et al., 2013 ; Diaz-Toribio & Putz, 2021 ).
Temuan kami menunjukkan bahwa batang semak Helichrysum yang hangus seluruhnya memiliki tunas ulang yang lebih banyak dibandingkan dengan batang yang terbakar sebagian dan tidak terbakar. Sementara penelitian saat ini menentukan kemampuan tunas ulang individu spesies Helichrysum yang terpengaruh oleh api secara berbeda, penelitian oleh Quevedo et al. ( 2011 ) menentukan efek rezim api pada spesies tanaman yang berbeda. Temuan dari penelitian sebelumnya mengungkapkan bahwa individu Acer sp. yang terbakar seluruhnya semuanya tumbuh kembali, sementara individu Juniperus communis dan Juniperus phoenicea yang terbakar kurang parah menunjukkan kemampuan tunas ulang yang rendah (Quevedo et al., 2007 ). Selain itu, temuan dari penelitian saat ini menunjukkan bahwa semak yang tidak terbakar menghasilkan lebih sedikit tunas ulang dibandingkan dengan semak yang terbakar seluruhnya dan sebagian, kemungkinan karena mereka mengalokasikan sumber daya mereka secara lebih merata di antara tunas lama yang ada dan tunas ulang baru (Clarke et al., 2013 ; Guo et al., 2022 ). Kemampuan tumbuh kembali yang kuat dari semak Helichrysum yang hangus sepenuhnya menunjukkan kemampuan mereka untuk pulih meskipun dalam kondisi kebakaran yang ekstrem. Respons kompensasi ini menunjukkan bahwa semak Helichrysum ini sangat beradaptasi dengan lingkungan yang rawan kebakaran, yang berpotensi mempersulit upaya untuk mengelola perluasannya. Respons pertumbuhan kompensasi Helichrysum setelah kebakaran intensitas tinggi dapat mendorong perkembangbiakan spesies ini melalui pertumbuhan kembali, yang menantang strategi pengelolaan yang dimaksudkan untuk membatasi penyebarannya di lingkungan yang rentan. Jumlah tunas kembali mencapai maksimum pada bulan Desember, hanya 3 bulan setelah kebakaran, yang mencerminkan kemampuan tumbuh kembali semak setelah kebakaran, tetapi kemudian menurun secara signifikan seiring berjalannya waktu. Pola pertumbuhan kembali ini mencerminkan tren regenerasi tunas kembali fakultatif di lingkungan yang rawan kebakaran (Underwood et al., 2023 ). Penurunan jumlah tunas baru secara bertahap seiring berjalannya waktu mungkin disebabkan oleh alokasi sumber daya ganda antara pembibitan dan pembentukan tunas baru, yang merupakan fenomena umum pada tunas baru fakultatif (Paula & Ojeda, 2006 ).
4.2 Dampak kebakaran terhadap regenerasi bank benih tanah spesies Helichrysum
Area yang terbakar akhir dan awal menunjukkan tingkat rekrutmen bibit yang rendah dibandingkan dengan area yang tidak terbakar. Temuan ini sejalan dengan penelitian yang dilakukan di wilayah lain (Cury et al., 2020 ; Lipoma et al., 2018 ; Shi et al., 2022 ), yang melaporkan bahwa lokasi yang tidak terbakar menunjukkan tingkat rekrutmen bibit tertinggi dari bank benih tanah, dengan penurunan rekrutmen pada kedalaman tanah yang lebih besar. Temuan ini menunjukkan bahwa benih Helichrysum tidak memiliki mekanisme perlindungan terhadap api, tidak seperti spesies semak lainnya seperti Erica coccinea L. subsp. Coccinea (Bell & Ojeda, 1999 ). Menurut Steger et al. ( 2022 ), tekstur kertas benih Helichrysum menunjukkan keberadaan utama mereka di tanah permukaan, membuatnya rentan terhadap kerusakan akibat kebakaran. Oleh karena itu, pembakaran akhir dapat secara efektif mengurangi perambahan Helichrysum karena sebagian besar benih Helichrysum rentan terhadap kerusakan akibat kebakaran.
4.3 Pengaruh pembakaran dini dan pembakaran akhir terhadap regenerasi Helichrysum melalui bibit di seluruh kedalaman tanah
Kami menemukan bahwa dampak kebakaran pada benih berkurang pada kedalaman tanah lebih dari 2 cm, yang mengonfirmasi bahwa suhu tanah kemungkinan menurun seiring bertambahnya kedalaman selama kebakaran (Afolayan et al., 1997 ; Williams et al., 2004 ; Xu et al., 2022 ). Studi lain tentang dampak kebakaran pada perkecambahan benih menunjukkan bahwa sementara tingkat panas antara 110°C mengurangi perkecambahan benih Daucus carota L, intensitas panas yang sama meningkatkan perkecambahan benih Helichrysum foetidum (L.) Moench dan Oenothera glazioviana Micheli (Riveiro et al., 2019 ).
Kami menyimpulkan bahwa kebakaran mungkin berdampak negatif pada perkecambahan benih dan pembentukan spesies Helichrysum di area yang terbakar awal dan akhir. Namun, kami juga menunjukkan bahwa tegakan Helichrysum yang mapan kemungkinan besar tahan terhadap kebakaran dan dapat bertunas kembali setelah kebakaran, berpotensi menghasilkan banyak batang baru, bahkan di lokasi yang terbakar akhir. Meskipun studi ini hanya mencakup periode waktu yang singkat setelah kebakaran, studi ini memberikan wawasan berharga tentang dampak kebakaran pada regenerasi Helichrysum , area yang sejauh ini menunjukkan studi empiris yang terbatas. Sebagian besar literatur yang ada tentang genus Helichrysum dan ekologi kebakarannya berfokus pada lingkungan yang terkendali (Afolayan et al., 1997 ; Giovannini et al., 2008 ; Makena et al., 2023 ; Mott, 1972 ; Riveiro et al., 2019 ), sementara studi kami mencakup observasi lapangan dan studi ex-situ. Temuan kami memberikan pemahaman lebih jauh tentang respons potensial fenologi spesies Helichrysum dan perluasan populasi karena spesies lain seperti Helichrysum petiolare dan H. foetidum telah dipicu oleh kebakaran hutan dalam perluasan mereka ke habitat baru (Prunera-Olivé et al., 2019 ). Temuan penelitian ini menggarisbawahi pembakaran api awal dan akhir sebagai salah satu alat manajemen ekologis yang dapat menghambat perambahan semak Helichrysum di ekosistem padang rumput.
5 KESIMPULAN
Kami menyimpulkan bahwa pembakaran dini dan akhir dapat menjadi pendekatan pengelolaan yang efektif untuk menghambat perluasan semak Helichrysum ke padang rumput, terutama karena perkecambahan biji yang rendah yang kami temukan saat terjadi kebakaran. Karena kami melaporkan jumlah biji Helichrysum tertinggi di permukaan tanah dibandingkan dengan lapisan tanah yang lebih dalam, kami menyatakan bahwa biji-biji ini akan paling rentan terhadap kerusakan akibat kebakaran. Kami mengusulkan bahwa pembakaran dini dan akhir mengurangi potensi perambahan semak Helichrysum jangka panjang karena kedua peristiwa pembakaran berdampak negatif pada perkecambahan biji Helichrysum . Namun, kami juga menunjukkan bahwa pembakaran dini maupun akhir mungkin tidak cukup untuk memusnahkan semak Helichrysum yang sudah tumbuh di TNK, karena kami menemukan semak tersebut cukup tahan terhadap kebakaran, terutama melalui kemampuan mereka untuk tumbuh kembali. Hal ini menunjukkan bahwa kebakaran saja mungkin tidak cukup sebagai strategi pengendalian jangka panjang untuk spesies semak ini. Oleh karena itu, kami mengusulkan bahwa strategi pengelolaan di masa mendatang harus mempertimbangkan waktu dan tahap fenologi tanaman Helichrysum yang merambah . Mengingat penelitian ini hanya mencakup periode 5 bulan pasca-kebakaran, kami merekomendasikan studi pemantauan jangka panjang untuk menyelidiki aspek kebakaran tambahan, seperti pengaruh frekuensi kebakaran terhadap kemampuan regenerasi Helichrysum dan semak lainnya di KNP.